Ahhoz, hogy átfogóan tudjunk beszélni a hemoglobinról (Hb), célszerű először kezelni a mioglobint (Mb), amely nagyon hasonlít a hemoglobinnal, de sokkal egyszerűbb. Szigorú rokonsági viszony áll fenn a hemoglobin és a myoglobin között: mindkettő konjugált fehérje, és protetikai csoportja (nem fehérje) a hem csoport.
A myoglobin egy gömbfehérje, amely egy-egy láncból áll, mintegy százötven aminosavból (a szervezettől függően) és molekulatömege körülbelül 18 Kd.
Amint már említettük, egy hem csoporttal van ellátva, amely a fehérje hidrofób (vagy lipofil) részébe van beillesztve, amely a rostos fehérjék α-hélix struktúráinak tulajdonítható hajtásokból áll.
A myoglobin főleg α-hélixek szegmenseiből áll, amelyek nyolcban vannak jelen, és szinte kizárólag nem poláros maradványokat (leucin, valin, metionin és fenilalanin) tartalmaznak, míg a poláris maradékok gyakorlatilag hiányoznak (aszparaginsav, glutaminsav, lizin és arginin); az egyetlen poláros maradék két hisztidin, amelyek alapvető szerepet játszanak az oxigén támadásában a hem csoporthoz.
A hem csoport a kromofór csoport (a láthatóban felszívódik) és a mioglobin funkcionális csoportja.
Lásd még: glikált hemoglobin - hemoglobin a vizeletben
Egy kis kémia
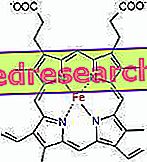
A protoporfirin és a vas közötti kapcsolat az úgynevezett koordinációs vegyületek tipikus kötése, amelyek olyan kémiai vegyületek, amelyekben egy központi atom (vagy ion) más vegyi anyagokkal kötődik, mint az oxidációs száma (elektromos töltés). A hem esetében ezek a kötések reverzibilisek és gyengék.
A vas koordinációs száma (koordinációs kötések száma) hat: a vas körül hat molekula lehet, amelyek megosztják a kötő elektronokat.
Koordinációs vegyület létrehozásához két helyes tájolású orbitumra van szükség: az elektronokat "vásárolni", a másik pedig adományozhatja őket
A hemban a vas négy sík kötést képez a proto-porfirin gyűrű közepén lévő négy nitrogénatommal és egy ötödik kötéssel egy proximális hisztidin nitrogénnel; a vas a hatodik szabad koordinációs összeköttetéssel rendelkezik, és az oxigénhez kötődik.
Amikor a vas szabad ion formájában van, d- típusú orbitáik mindegyike ugyanolyan energiával rendelkezik; a mioglobinban a vasion kötődik a protoporfirinhez és a hisztidinhez: ezek a fajok mágnesesen zavarják a vas pályáit; a perturbáció mértéke a különböző orbitákban térbeli orientációjuk és zavaró fajok szerint eltérő lesz. Mivel a pályák teljes energiájának állandónak kell lennie, a perturbáció energikus szétválasztást eredményez a különböző orbiták között: az egyes orbiták által megszerzett energia egyenértékű a többi elveszett energiával.
Ha az orbitálisok közötti szétválasztás nem túl nagy, akkor előnyös a magas spin elektronikus elrendezés: a kötő elektronok párhuzamosan forognak több lehetséges alszintben (maximum multiplicitás); másrészt, ha a perturbáció nagyon erős, és nagy az elválasztás az orbiták között, célszerűbb a kötőelektronok párosítása az alacsonyabb energiájú orbitákban (alacsony centrifugálás).
Amikor a vas oxigénhez kötődik, a molekula alacsony centrifugálással rendelkezik, míg amikor a vas a hatodik szabad koordinációs összeköttetéssel rendelkezik, a molekula magas centrifugálással rendelkezik.
Ennek a spin-különbségnek köszönhetően a mioglobin spektrális analízisén keresztül meg tudjuk érteni, hogy van-e kötődő oxigén (MbO2) vagy nem (Mb).
A myoglobin az izmokra jellemző fehérje (de nem csak az izmokban található).
A myoglobint a spermiumbálából nyerik ki, amelyben nagy mennyiségben van jelen, majd tisztítjuk.
A cetfélék lélegeztetnek, mint az embereké: a tüdőt el kell felszívniuk a légutakon keresztül; a spermium bálának annyi oxigént kell hordoznia az izmokban, amelyek képesek oxigént felhalmozódni azáltal, hogy kötődnek az általuk jelen lévő myoglobinhoz; az oxigént ezután lassan szabadítják fel, amikor a cetfélék elmerülnek, mert az anyagcsere oxigént igényel: minél nagyobb az oxigénmennyiség, amit a spermium bálna képes felszívni, és minél több oxigén áll rendelkezésre a merülés során.

Ahhoz, hogy a hús többé-kevésbé piros legyen, a hemoproteinek tartalma (ez a hém, amely a húst pirosá teszi).
A hemoglobin sok szerkezeti analógiával rendelkezik a myoglobinnal és képes a molekuláris oxigént reverzibilisen kötni; de míg a myoglobin általában az izmokra és a perifériás szövetekre korlátozódik, a hemoglobin az eritrocitákban vagy a vörösvérsejtekben található (ezek pszeudo-sejtek, azaz nem valós sejtek), amelyek a vér 40% -át teszik ki.
A hemoglobin tetraméter, azaz négy polipeptidláncból áll, mindegyikben egy hem csoport és két kettő (egy emberben két alfa-lánc és két béta lánc).
A hemoglobin fő funkciója az oxigén szállítása ; a vér másik funkciója, amelyben a hemoglobin részt vesz, az anyagok szövetekre való szállítása.
A tüdőből (oxigénben gazdag) a szövetekbe vezető úton a hemoglobin oxigént hordoz (ugyanakkor más anyagokat is eljut a szövetekhez), míg a fordított úton a szövetekből gyűjtött hulladékot, különösen az anyagcsere során keletkező szén-dioxidot hordozza.
Az ember fejlődésében olyan gének vannak, amelyek csak egy bizonyos ideig fejeződnek ki; emiatt különböző hemoglobinok vannak: a felnőtt ember magzati, embrionális.
A különböző hemoglobinokat alkotó láncok különböző szerkezetekkel rendelkeznek, de hasonlóságukkal az általuk végzett funkció többé-kevésbé azonos.
A különböző láncok jelenlétének magyarázata a következő: a szervezetek evolúciós folyamata során a hemoglobin is fejlődött az oxigén szállítására specializálódott területeken, ahol hiányos területek vannak. Az evolúciós lánc elején a hemoglobin kis organizmusokban oxigént hordozott; az evolúció során a szervezetek nagyobb méreteket értek el, ezért a hemoglobin megváltozott, hogy képes legyen oxigént szállítani azon területekre, amelyek gazdagabbak; Ennek érdekében az evolúciós folyamat során a hemoglobint alkotó láncok új struktúráit kodifikálták.
A myoglobin még mérsékelt nyomáson is kötődik az oxigénhez; perifériás szövetekben körülbelül 30 mmHg nyomás van (PO2): ezen a nyomáson a myoglobin nem szabadít fel oxigént, így az oxigén hordozóként hatástalan lenne. A hemoglobin ugyanakkor rugalmasabb viselkedést mutat: magas nyomású oxigént köt, és a nyomás csökkenésekor felszabadítja azt.
Ha egy fehérje funkcionálisan aktív, enyhén megváltoztathatja alakját; például az oxigénezett myoglobin más formában van, mint a nem oxigénezett myoglobin, és ez a mutáció nem befolyásolja a szomszédosakat.
A diskurzus különbözik a kapcsolódó fehérjék, például a hemoglobin esetében: amikor egy lánc oxigénnel van ellátva, úgy alakul ki, hogy megváltoztatja alakját, de ez a módosítás háromdimenziós, így a tetraméter többi láncát is befolyásolják. Az a tény, hogy a láncok egymáshoz kapcsolódnak, azt feltételezi, hogy az egyik módosítása más szomszédokat is érint, még ha másképpen is; amikor egy lánc oxigénnel van ellátva, a tetraméter más láncai "kevésbé ellenséges hozzáállást" vesznek fel az oxigén felé: az a nehézség, amellyel a lánc oxigenátjai csökkennek, mint a mellette lévő láncok oxigenizálnak. Ugyanez az érv érvényes az oxigénmentesítésre.
A deoxihemoglobin kvaterner struktúráját T (feszült) formának nevezzük, míg az oxihemoglobin R formáját (szabaddá) nevezik; a feszített állapotban a sav aminosavak és a bázikus aminosavak között meglehetősen erős elektrosztatikus kölcsönhatások sorozata van, amelyek a deoxihemoglobin merev szerkezetéhez vezetnek (ezért a "feszült forma"), miközben az oxigén kötődik, ezek egysége az interakciók csökkennek (ezért a "kiadott forma"). Továbbá oxigén hiányában a hisztidin töltést (lásd a szerkezetet) az aszparaginsav ellentétes töltése stabilizálja, míg oxigén jelenlétében a fehérje hajlamos egy protont elveszíteni; mindez azt jelenti, hogy az oxigenált hemoglobin erősebb sav, mint a deoxygenált hemoglobia: bohr hatás .
A pH-tól függően a hem csoport többé-kevésbé könnyen kötődik az oxigénhez: savas környezetben a hemoglobin könnyebben szabadul fel az oxigénből (a feszült forma stabil), míg az alapvető környezetben az oxigénkötés az oxigénnel. erősebb.

Minden hemoglobin 0, 7 protont bocsát ki bejövő mól móljára (O2).
A Bohr hatás lehetővé teszi, hogy a hemoglobin javítsa az oxigén hordozhatóságát.
A hemoglobinnak, amely a tüdőből a szövetekbe utazik, kiegyensúlyoznia kell a nyomást, a pH-t és a hőmérsékletet.
Látjuk a hőmérséklet hatását .
A pulmonáris alveolák hőmérséklete 1-1, 5 ° C-kal alacsonyabb, mint a külső hőmérséklet, míg az izmokban a hőmérséklet körülbelül 36, 5-37 ° C; a hőmérséklet emelkedésével a telítési tényező csökken (ugyanazzal a nyomással): ez azért történik, mert a kinetikus energia növekszik és a disszociáció kedvező.

Vannak más tényezők is, amelyek befolyásolhatják a hemoglobin oxigénhez való kötődésének képességét, amelyek közül az egyik a 2, 3-biszfoszfoglicerát koncentrációja .
A 2, 3-biszfoszfoglicerát egy 4-5 mM koncentrációjú eritrocitákban jelenlévő metabolikus szer (a test egyik más részében ilyen magas koncentrációban nincs jelen).
Fiziológiai pH-nál a 2, 3-biszfoszfoglicerát deprotonálódik és öt negatív töltéssel rendelkezik; ez a hemoglobin két béta-láncának ékébe megy, mert ezek a láncok magas koncentrációjú pozitív töltésekkel rendelkeznek. A béta-láncok és a biszfoszfoglicerát láncok közötti elektrosztatikus kölcsönhatások bizonyos merevséget adnak a rendszernek: feszült szerkezetet kapunk, amely kevés affinitással rendelkezik az oxigénnel; az oxigénellátás során a 2, 3-biszfoszfoglicerátot eltávolítjuk.
Az eritrocitákban van egy speciális berendezés, amely átalakítja az 1, 3-biszfoszfoglicerátot (az anyagcsere által termelt) 2, 3-biszfoszfogliceráttá, így eléri a 4-5 mM koncentrációt, és így a hemoglobin képes kicserélni a l oxigén a szövetekben.
A szövetbe érkező hemoglobin a felszabadult állapotban van (oxigénhez kötődik), de a szövet közelében karboxilálódik, és egy kinyújtott állapotba jut: az ebben az állapotban lévő fehérje alacsonyabb hajlamot köt az oxigénnel, a felszabadult állapothoz képest a hemoglobin oxigént szabadít fel a szövetbe; továbbá a víz és a szén-dioxid közötti reakció miatt H + ionok képződnek, ezért a bohr hatás miatt további oxigén keletkezik.
A szén-dioxid diffundál a plazmamembránon áthaladó vörösvértestbe; mivel a vörösvértestek a vér mintegy 40% -át teszik ki, azt várjuk, hogy a szövetekből diffundáló szén-dioxid 40% -a belép belőle, a valóságban a szén-dioxid 90% -a belép a vörösvértestekbe, mert olyan enzimet tartalmaznak, amely átalakítja a \ t szén-dioxid szén-dioxidban következik, hogy a szén-dioxid állandós koncentrációja az eritrocitákban alacsony, ezért a belépés sebessége magas.

Egy másik jelenség, amely akkor következik be, amikor az eritrocita eléri a szövetet, a következő: gradiens szerint a HCO3- (szén-dioxid-származék) jön ki az eritrocitából, és a negatív töltés kilépésének kiegyensúlyozásához a következő: kloridok belépése, amelyek meghatározzák az ozmotikus nyomás növekedését: a változás kiegyensúlyozása érdekében a víz is belép, amely eritrocita duzzanatot okoz (HAMBURGER hatás). Az ellenkező jelenség akkor fordul elő, amikor az eritrocita eléri a pulmonáris alveolákat: az eritrocita defláció lép fel (HALDANE hatás). Így a vénás eritrociták (közvetlenül a tüdőbe) gyengébbek, mint az artériásak.